
9. X-Pigeons |
||||
You are here because you want to understand the colors and patterns of your pigeons, and breed your desired offspring. You would like to know which parents you need to breed together to get that desired trait. Perhaps you are wondering why you can’t get a black and white grizzle when you mate a white-self bird with a black-self bird. You want to understand how you got a blue youngster from two red parents. You are wondering if you could get an almond offspring when you breed a yellow bird with a brown bird, then breed the offspring from this mating with the offspring of black and white parents.
Ancient people were also concerned with genetics and inheritance and sometimes developed some fantastic ideas about it because they did not understand our present concept of genetic science. They believed in mermaids, a mythological aquatic creature with a female human head and torso and the tail of a fish. Another example of the ancients' fascination with hybrid offspring is Pegasus -- a winged horse in Greek mythology, representing a divine horse that was fathered by Poseidon. In the ancient Arabic culture, it was believed that if you crossed a camel with a sparrow, you would get an ostrich, or you would get a wild boar if you crossed a goat with a dog. We can get pouter pigeons when we cross a rooster with a bull frog. Then cross the offspring to a homing pigeon. Of course, you see in the picture below a prime example of what happens when you breed a pony with a Birmingham Roller.
 Unfortunately,
genetic science can be a very complicated subject. It is even harder
to write about genetics because there are still many pigeon colors
and patterns, the inheritance of which is almost entirely unknown.
The fact is, no one really knows all the genetic parts needed to make
a human being. Hence, not much is known about pigeon genetics. Not
only do we not know what all the genes are and where they are located,
we don't even know how many there are. Therefore, like the rest of
genetics, pigeon genetics still has a lot of unknown elements.
Unfortunately,
genetic science can be a very complicated subject. It is even harder
to write about genetics because there are still many pigeon colors
and patterns, the inheritance of which is almost entirely unknown.
The fact is, no one really knows all the genetic parts needed to make
a human being. Hence, not much is known about pigeon genetics. Not
only do we not know what all the genes are and where they are located,
we don't even know how many there are. Therefore, like the rest of
genetics, pigeon genetics still has a lot of unknown elements.
So far, we only know about 50 different mutants in pigeons in detailed behavior, but we know that there are many, many more that we do not understand in detail yet. Nonetheless, science moves forward swiftly. We live in a time when if you do not keep up with these rapid changes, what you think you know has simply been replaced by new discoveries much of the time. What is known as classical genetics, which is a field concerned with the explanation of traits and how they are inherited from parents to offspring, has been updated with the advent of molecular biology and biochemistry.
Attacking the genetic problems at the biochemical level after the Hollander Era has already started to pay off for pigeon genetics. With the start of the Genomic Era in pigeon genetics, results found in testing tubes in labs are already proving to be much more accurate than conventional Mendelian results. Thus, if you learned about genetics in the '60s or '70s and did not keep up, you are so hopelessly behind that what you know is next to useless beyond the most rudimentary problems. However, although the advent of yeast molecular genetics has introduced new and exciting capabilities, classical genetics still remains the approach of choice in many instances.
In any case, if you are a beginner and have no prior knowledge in genetics, don’t panic yet. I intend to cover the basic genetics first. The information in these pages is not a comprehensive work intended to cover all aspects of pigeon genetics, but instead helps pigeon breeders understand how traits are inherited and how this can help them breed better quality of pigeons. If you are reading this page or pages like these, you are also one of those elite pigeon breeders with the right attitude who do not say, “This is a hobby, not a science project!” I am a computer science teacher, and biology has always been one of my favorite subjects. I am neither a scientist nor a geneticist. I am just a curious, albeit serious, pigeon breeder, and I would like to understand as much as I can about my hobby to produce the desired offspring. Although I have been experimenting with genetics since 2007, most of the information in this website comes from research I have done using Internet forums and email groups, books, science magazines, and my interaction with other elite pigeon breeders and, most importantly, with scientists.
I would like to give special thanks and appreciation to Dr. Richard Cryberg (chemist by training) for his help and his contributions to this website and for assisting me in every aspect of pigeon genetics. Dr. Cryberg's exceptional knowledge in genetics and chemistry, his patience and willingness to assist, where possible, is greatly appreciated. I would also like to thank Mr. Frank Mosca (biologist by training), who also helped me to understand the basics of pigeon genetics and allowed me to use his website as one of the references. Without these two great scientists of the hobby mentioned above, I would still be able to study and write about basic pigeon genetics, but I highly doubt I could learn it this fast with accuracy and with this kind of sophistication. My goal is to collect and report what we currently know about pigeon genetics so that future students of pigeon genetics after Hollander, Miller, and Quinn could start studying genetics with the most up-to-date information. I have also used a variety of references. Click here to view work cited.
 I
update this website very often as I gain more knowledge in pigeon genetics.
Although I try not to make it too difficult and technical,
if you are a beginner or have no prior knowledge in genetics, you might
want to spend some time reading about basic genetics on this page before
moving on to the next pages. Understanding the information, especially
about basic genetics, will take you above and beyond the many pigeon
breeders and fanciers. So don’t be discouraged and think it is
beyond your understanding. Although genetics can be very complicated,
it can also be a lot more interesting and a lot more fun than you might
think. In order to better understand pigeon genetics, you should be
familiar with basic genetics and genetic terms first. Thus, I will
start with some simple definitions, which will hopefully create some
familiarity with terms. When you get comfortable with the basic terms,
then the subject of pigeon genetics should not seem so difficult.
I
update this website very often as I gain more knowledge in pigeon genetics.
Although I try not to make it too difficult and technical,
if you are a beginner or have no prior knowledge in genetics, you might
want to spend some time reading about basic genetics on this page before
moving on to the next pages. Understanding the information, especially
about basic genetics, will take you above and beyond the many pigeon
breeders and fanciers. So don’t be discouraged and think it is
beyond your understanding. Although genetics can be very complicated,
it can also be a lot more interesting and a lot more fun than you might
think. In order to better understand pigeon genetics, you should be
familiar with basic genetics and genetic terms first. Thus, I will
start with some simple definitions, which will hopefully create some
familiarity with terms. When you get comfortable with the basic terms,
then the subject of pigeon genetics should not seem so difficult.
So, are you ready to explore pigeon genetics? All right! This is mission control. For your safety, please buckle up, remain seated, and keep your eyes on the monitor at all times. No talking, sleeping, smoking, eating, drinking, videotaping, flash photography, copying, or pasting while you are visiting this website. These rules are for the safety of the readers to avoid the severe damages to themselves as well as the safety of their domestic pigeon breeds. Mumtaztic Loft will not be responsible for any liability. I highly recommend that before you are cleared for launch, read the following friendly warning if you would like to survive pigeon genetics.
For those of you who might be thinking about breaking the rules above, I have genetically engineered the best loft protector, scientific name "Columba Raptor," to help me keep the order around here. For your information, the disobedient readers will be penalized accordingly. Let me tell you what happened to the last guy who attempted to steal the copyrighted material form this website without my permission.
When he thought about highlighting a paragraph, his lungs punctured in a matter of seconds, but his death did not come without pain. While he was screaming for help, hanging in the air on the raptor’s talons, flying back to his nest, the perpetrator's head was gently but slowly separated from his body and used as a snack for the remainder of the flight.
The rest of his useless body was fed to the little raptors who were impatiently waiting for their parents to bring food to them. Columba Raptors are capable of carrying weights up to three hundred pounds (137 kg). The blue checks are usually faster and deadlier than the ash-red bars. They can smell and hear ignorance from miles away, and they are specifically bred to seek and destroy ignorant people. I suggest you act like you understand what you are about to read, and try to look calm at all times. Thank you for following the rules and not violating any copyright laws; I hope you will enjoy the rest of this website. You are now cleared for launch. Travel at your own risk!
All right, let’s get serious and start learning genetics, which is not as deadly as the Columba Raptor which is created by Adobe Photoshop (a raster graphics editing software), and not by nature. Let me start with the brief history of life on Earth to give you an idea how long it took to get this far.
Approximately 4.6 billion years ago, Earth was formed. We know this because radioactive dating of rocks found on the moon, meteorites, and Earth all give this as the maximum date. In its 4.6 billion years circling the sun, the Earth has harbored an increasing diversity of life forms, but it wasn’t until 4.0 billion years ago that Earth started to cool down to became viable for living things to exist on it. Conditions on Earth four billion years ago were very different than they are today. The atmosphere lacked oxygen, and an ozone layer did not yet protect Earth from harmful radiation. Heavy rains, lightning and volcanic activity were common. Yet the earliest cells originated in this extreme environment. Today, a group of single-celled organisms called archaeabacteria, or archaea, still thrive in extreme habitats. We don’t know how life originated; it probably arose on Earth, but an alternate theory suggests it arose elsewhere and drifted through space to seed the Earth. Many theories of the origin of life have been proposed, but since it's hard to prove or disprove them, no fully accepted theory exists. About 3.7 billion years ago, the first prokaryotic (single cell) organism was present on Earth when there was no oxygen in the atmosphere. About 3.4 billion years ago, cyanobacteria started to perform photosynthesis. It took almost another 2 billion years—roughly 1.5 billion years ago—in which nucleated cells through nucleus evolved. Thus, it took 300 million years to evolve life on Earth, but it took another 2 billion years to be able to evolve the nucleus. It seems that evolving a nucleus was a big deal and hard to do, and likewise, evolving past single cells was also a big deal and hard to do. Both of these steps seem harder than it was to form the first life, based on the amount of time it took for each step. Perhaps it was because first cells didn’t have any competition to survive. About a half billion years ago, eukaryotic (multicellular) life evolved. Hence, these dates suggests that making life start was not all that hard, but making life evolve into something complex was a major step for life on Earth. About 600 million years ago, simple animals started to evolve. Five hundred million years ago, fish and proto-amphibians; 475 million years ago, land plants; 400 million years ago, insects and seeds; 360 million years ago, amphibians; 300 million years ago, reptiles; 200 million years ago, mammals; 150 million years ago, birds; 130 million years ago, flowers; 60 million years ago, the primates, 20 million years ago, the family Hominidae (great apes); 2.5 million years ago, the genus Homo (including humans and their predecessors); 200,000 years ago, homo sapiens (anatomically modern humans) evolved. Today, the science proves that about 98% of human and chimp DNA are the same, but that does not mean humans evolved from apes. It simply means that like all life forms on Earth, apes and humans share a common ancestor that lived millions of years ago.
Now that I covered the summary of life on Earth, let’s start with the basic genetic terms and concepts. But before I start, let me share some quotes from two of the major contributors and most respected scientists of pigeon and dove genetics.
“Genetics is the study of the nature and mechanism of inheritance. But as a definition it leaves much to be desired. A better characterization describes its aims: to develop principles and methods whereby we can make predictions concerning the expression and transmission of similarities and dissimilarities between individuals and populations of organism” (Dr. Wilmer J. Miller, A Survey of Genetics).
We cannot say that expertise in Genetics is essential to success in showing, racing, or even squab farming. Some understanding however can be helpful and gratifying. But like all science, Genetics is never finished: the pigeons will continue to give us mysteries for solutions. Any fancier may discover something that is beyond our knowledge, and the fancier may even discover the answer. Generally we are not adequately prepared to pursue the myriad problems that arise, but perhaps collaboration will develop. Pigeon Genetics Newsletter, and its successor, Pigeon Science and Genetics Newsletter, have been leading the way, trying to keep up communication. Other periodicals have also been helpful (Dr. Willard F. Hollander, Origins and Excursions in Pigeon Genetics, P.3).
Genetics,
the science of genes, heredity, and the variation of organisms, is
not like most subjects in biology. Knowledge of genetics is fundamental
to an understanding of how organisms, populations, and species evolve,
behave, and receive common traits from previous generations. An important
part of genetics is problem-based, and it requires us to be able to
organize information, draw conclusions from that information, and find
solutions to problems. Because of this, reading and learning genetics
is more like studying Algebra and less like studying anatomy. Therefore,
a basic understanding of genetics is required in order to comprehend
how the multitude of pigeon varieties came to exist and how their traits
are inherited. An understanding of basic cytogenetics is required before
proceeding into specific pigeon mutation crosses and the expected outcome.
Cytogenetics is the science which attempts
to correlate cellular events with genetic phenomena. 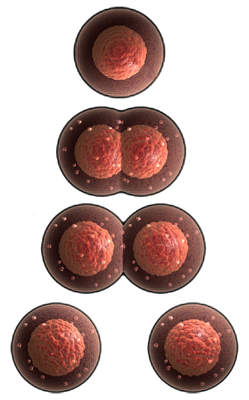
The Cell
The Earth is teeming with countless living cells. We are all made out of cells, and the human body contains about 100 trillion cells. A cell is the smallest, most fundamental unit of life. Cells provide structure for the body, take in nutrients from food, convert those nutrients into energy, and carry out specialized functions. Cells also contain the body’s hereditary material and can make copies of themselves. Cell numbers increase by cell division, where a mother cell divides into two daughter cells. During cell division, it is extremely important that the daughter cells inherit the correct and complete information from their mother cell. Cell division is the essential feature of life, and we should look at cell division as almost the definition of life. All cells are descendants (offspring) from cells that have divided, and cells must have divided to produce ancestors (parents). The ancestry of living things extends backwards to their parents, and their parents’ parents, etc. Our descendants, assuming we have any, will exist in future generations.
Life on Earth is categorized by unicellular and multicellular organisms. Some organisms are nothing more than a single cell for their life span, so we call them unicellular organisms. These organisms make up most of the life on Earth. Unicellular organisms are microscopic and require microscopes for viewing, so many of us never see them. Unicellular organisms are a primitive group of cells with no complex organization or specialization, such as bacteria. They are single-celled organisms, wherein the functions like feeding, locomotion, expelling wastes, reproduction, etc. are carried out by the single cell. Each of the unicellular organisms possesses specific characteristic features, which help in adaptation to a wide range of environmental conditions, and some of them divide more than once every hour. Unicellular organisms are as diverse as they are ubiquitous; you can find unicellular organisms in every habitat, even in the least hospitable environments—from the deep ocean to the edges of hot springs to just about every surface of our bodies. The main groups of unicellular organisms are bacteria, archaea, protozoa, unicellular algae, and unicellular fungi. Unicellular organisms fall into two general categories: prokaryotic organisms — lacking a membrane-bound nucleus (karyon), mitochondria or any other membrane-bound organelles and eukaryotic organisms — containing a nucleus and other structures like organelles. It is believed that the oldest forms of life, unicellular organisms, existed about 3.7 billion years ago.
Organisms such as you and I are a collection of many cells, so we are called multicellular organisms. It is important to understand, however, all multicellular organisms start as a single cell. We become multicellular by a series of cell divisions very similar to the way unicellular organisms reproduce, but instead of drifting apart as two separate cells, most of our cells stick together and eventually organize themselves into a specialized "colony" that grows into a multicellular organism. We do not know what triggered the transition from single-cells to multicellular life, how the strategies arose that support today's multicellular organisms, or what alternatives were tried and failed. It seems likely that an early step in the evolution of multicellular organisms was the association of unicellular organisms to form colonies. The simplest way of achieving this is for daughter cells to remain together after each cell division. By collaboration and by division of labor, it becomes possible to exploit resources that no single cell could utilize so well. Even though we like to divide species into unicellular and multicellular organisms, it is not entirely correct because at one time we were also only one cell -- but we outgrew it after the sperm cell from our father fertilized the egg cell in the womb of our mother (conception). We went from a single cell to a multicellular organism. Cell division, especially in multicellular organisms, is under very careful control. From the fertilization of the egg to the birth of a child, cells divide to make a multicellular organism. If the cells of an unborn baby underwent just one extra cell division, that baby would be twice as big, and I wouldn’t want to imagine a female giving birth to that baby.
 Gene
Gene
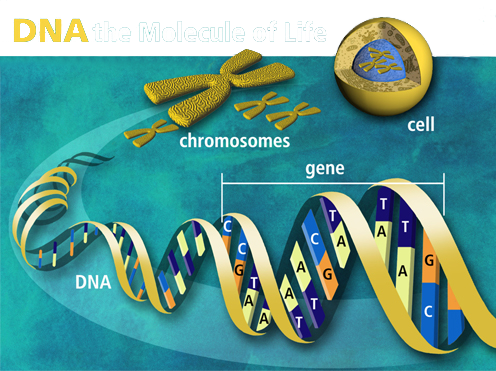
The instructions to build a cell are coded as genetic information, and we call them genes, located inside the nucleus of the cells. Genes are a set of instructions that decide what the organism is like, how it survives, and how it behaves in its environment. Genes are also the basic unit of heredity, capable of transmitting characteristics from one generation to the next. Hence, genes carry information that determines each person’s traits. Traits are characteristics that organisms inherit from their parents, such as the color of their hair, the color of their skin, the color of their eyes, height, likelihood to develop certain diseases, etc.
Genes can act together to produce a single trait, and we know that at least four genes control height in humans. Skin color is determined by at least three genes in humans. It seems evident that traits like rolling/tumbling and color mutations like Toy Stencil, Frill Stencil are caused by multiple genes in pigeons. Sometimes, the effects of genes are altered by the environment. For instance, if your diet is lacking in protein, minerals, and vitamins, you won’t grow as tall. Humans today are 10 cm taller than they were in the 1800s.
So, all living things have genes, and all human beings have the same set of genes since we are all of the same species, but we don’t have the same versions of genes. Thus, we all look different from one another unless we have an identical twin.
All genes that are present and working to perform their normal function are called wild-type genes. Any gene that is not performing normally because something happened to it is called a mutant gene. The words wild-type originally were conceptualized as a product of the standard "normal" in contrast to that produced by a non-standard "mutant". The concept of wild-type is especially important to understand because all of our domesticated pigeons are mutants. I will cover wild-type and mutation subjects in more detail in the following pages.
Until we discovered genes, we thought we inherited factors from our parents and passed them to our offspring by blood. Aristotle (384-322 B.C) specified that the female contributed the building material of the progeny, and the male imparted the life force. This life force or vitalizing principle was suggested to stem from “purified” blood. This rather vague theory then became associated with blood and the pooling of blood of parents. The phrases like blood-line, blood-relation, pure-blooded are so embedded in society, they are still commonly used to identify a family or lineage with a common ancestry or the descendants of one individual. We now know that genes are the directions for building all the proteins that make our body function. All the genes in our body are needed to give instructions for how to make and operate all parts of our bodies. For example, blood contains red blood cells that transport oxygen around our bodies. The cells use a protein called hemoglobin to capture and carry the oxygen. If our hemoglobin gene is normal, the hemoglobin protein works fine. But if the instructions in that gene are changed or "mutated," changes in the hemoglobin protein could result. One such mutation causes a disorder called sickle cell anemia. Genes contain instructions for building proteins, which are involved in all sorts of things. Hemoglobin protein is just one example. Of our 25,000 genes, only a few contain the instructions for making hemoglobin proteins. The remaining genes contain the instructions for making other parts of our bodies. Other proteins, such as the enzymes that produce pigment in your eyes, and keratin, responsible for growing hair and nails, are also produced by genes.
Chromosome
 In DNA,
genes are all lined up on thread-like things called chromosomes which
come in pairs, and there are hundreds and sometimes thousands
of genes in one chromosome. Normally, each gene (except what is known
as the jumping genes) is located on a specific place on a chromosome.
Chromosome is a molecule containing genes found in the nucleus of a
cell. The word chromosome comes from the German, ultimately of Greek
roots chromo (color) + soma (body). The two partners known as the homologous
chromosomes, are similar in size, shape, and the position of their
centromeres. The 46 chromosomes in human cells constitute 23 homologous
pairs. The most important feature of homologous cells is that they
carry very similar, but not necessarily identical, genetic information.
One member of each homologous pair is called the maternal
homologue because it was originally inherited from the female parent; the other
member of a homologous pair is the paternal homologue because it was
inherited from the male parent. That's why children look like both
their mom and dad. Every individual of a given species has a characteristic
number of chromosomes. How many chromosomes does a cell hold? The correct
answer to this question depends on whether you're a fish or a fly or
a human. Chromosomes vary widely between different organisms, and they
do not correlate with the complexity of the organism. It is not the
number of chromosomes that makes each species unique but the information
the gene specifies.
In DNA,
genes are all lined up on thread-like things called chromosomes which
come in pairs, and there are hundreds and sometimes thousands
of genes in one chromosome. Normally, each gene (except what is known
as the jumping genes) is located on a specific place on a chromosome.
Chromosome is a molecule containing genes found in the nucleus of a
cell. The word chromosome comes from the German, ultimately of Greek
roots chromo (color) + soma (body). The two partners known as the homologous
chromosomes, are similar in size, shape, and the position of their
centromeres. The 46 chromosomes in human cells constitute 23 homologous
pairs. The most important feature of homologous cells is that they
carry very similar, but not necessarily identical, genetic information.
One member of each homologous pair is called the maternal
homologue because it was originally inherited from the female parent; the other
member of a homologous pair is the paternal homologue because it was
inherited from the male parent. That's why children look like both
their mom and dad. Every individual of a given species has a characteristic
number of chromosomes. How many chromosomes does a cell hold? The correct
answer to this question depends on whether you're a fish or a fly or
a human. Chromosomes vary widely between different organisms, and they
do not correlate with the complexity of the organism. It is not the
number of chromosomes that makes each species unique but the information
the gene specifies.
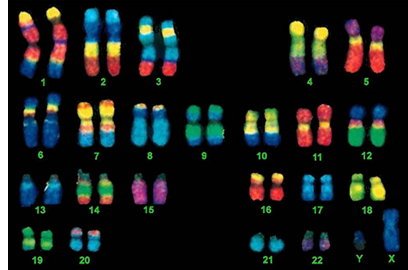
Humans are not unique in having 46 chromosomes; some other species - the olive tree, for example - also have 46 chromosomes. Humans have 23 pairs of chromosomes. Birds tend to have a rather large numbers of pairs, often about 50. Chickens and dogs have 39 pairs; alligators have 16 pairs; mosquitos have 3 pairs; onions have 8 pairs. There is an ant in Australia with only a single pair. A fern holds the record for the most chromosomes – more than 1200. A certain species of roundworm has only 2 chromosomes in each cell, whereas some crabs have as many as 200. Most animal and plant species have between 8 and 50 chromosomes per body cell. Numbers above and below these are uncommon. The picture above shows the 23 pairs of human chromosomes. In the picture, the sex chromosomes are labeled "X" and "Y." The sets of chromosomes in this picture are from a male; you can tell because females do not have a Y chromosome. Instead, they have two X chromosomes. We will talk about sex chromosomes and sex-linked genes shortly. The gene numbers also vary in different organisms, and the chromosome does not tell much at all about how many genes an animal should have. No one really knows all the genetic parts needed to make a human being. Most estimates place the human gene count in the neighborhood of 25,000 genes, which falls between the number of genes in a chicken and the number in a grape. Grape plants have 30,434 genes by the latest count. Chickens have 16,736 genes.
DNA
Ugh . . . DNA—another big word that is used often but not understood by many. The chromosomes and genes are made of DNA, which is short for deoxyribonucleic acid. DNA is the fundamental building block for an individual's entire genetic makeup. It is a component of virtually every cell in any multicellular organism’s body. DNA molecules are the long-term storage of information that is often compared to a set of blueprints or a code since it contains the instructions needed to construct other components of cells. DNA is in every cell of our body except for red blood cells (they do not have a nucleus). However, our blood has DNA because of white blood cells. If all the DNA from just one of our cells was stretched out (decondensed) and assembled into a single line, it would be almost two meters (about 7 feet) long. This immense amount of DNA information is contained in a space that is only 2 millionths of a millimeter thick. The smallest gene in humans has 850 of the bases, and some genes have more than a million bases.
The structure of the DNA molecule itself is identical among all living things, from an amoeba to a 150-ton whale, from a blade of grass to a redwood tree. It consists of sugar, phosphate, and four nitrogen bases: adenine (A), thymine (T), guanine (G), and cytosine (C). These four types of molecules which are called bases are linked by hydrogen bonds between the complementary base pairs in a very specific way: A always with T, and G always with C. The DNA of humans is composed of approximately 3 billion base pairs. It is amazing that this molecule, consisting of only six basic components (four bases, a phosphate, and a sugar) can contain all the information required to make almost a million types of animals and nearly a half-million species of plants.
The order or sequence of these bases determines the information available for building and maintaining an organism, similar to the way in which letters of the alphabet appear in a certain order to form words and sentences. Unlike the 26 letters in the English alphabet, DNA only has 4 letters and each word is only 3 letters long. Therefore, the possible number of unique three-letter words DNA can make is only sixty-four. But DNA's store of information comprises more than just four bases arranged into sixty-four different codons, a sequences of three nucleotides; just as the words of a language can be put together to form an infinite number of texts, so the codons on the DNA molecule can be ordered in innumerable ways. The four letters in DNA, A, G, C, and T, are strung together in pairs in specific combinations and lengths to spell our genes. This genetic code defines how these sequences specify which amino acid will be added next during protein synthesis.
 Let's
continue with the metaphor that DNA is a language. Unlike the English
language, instead of starting the first word with a capital letter
at the beginning of each sentence, and instead of ending that sentence
with a period, DNA has a code for starting the sentence (equivalent
of a capital letter), and it has a code for ending the sentence
(equivalent of a period). The DNA words are always three letters
long. There are no spaces between words and no punctuation in the
language, except for start and stop signals. It looks something
like this: ATGCTGACCTAAACTGTGCACTTGCGA.
Let's
continue with the metaphor that DNA is a language. Unlike the English
language, instead of starting the first word with a capital letter
at the beginning of each sentence, and instead of ending that sentence
with a period, DNA has a code for starting the sentence (equivalent
of a capital letter), and it has a code for ending the sentence
(equivalent of a period). The DNA words are always three letters
long. There are no spaces between words and no punctuation in the
language, except for start and stop signals. It looks something
like this: ATGCTGACCTAAACTGTGCACTTGCGA.
Let’s use an analogy to better understand information about our genes, chromosomes, and DNA. Assume we have a bookshelf with only 2 rows. Each row has 23 volumes of encyclopedias; so, we have a total of 46 volumes. Now, imagine each volume as one of our chromosomes. Each page of each book contains a unique set of sentences approximately 10 to 30 per page. In this analogy, the groups of codons that have been arranged in grammatically correct sentences to form specific proteins are called "genes." The nucleotide bases—A, T, G, or C— are the letters that make up each sentence on each page of the books. Each sentence is made up of a very specific sequence of letters. Each word is a specific protein (amino acid sequences) also known as codons. The entire set of the books is the DNA.
An important property of DNA is that it can self-replicate, or make copies of itself. Each strand of DNA in the double helix can serve as a pattern for duplicating the sequence of bases. This is critical when cells divide because each new cell needs to have an exact copy of the DNA present in the old cell.
DNA Replication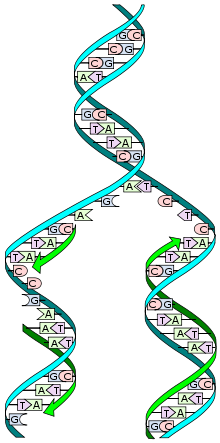
Each time a cell divides into two new cells, genes are copied, and the process that copies DNA is called DNA replication. Because the two strands of a DNA molecule have complementary base pairs, the nucleotide sequence of each strand automatically supplies the information needed to produce its partner. In other words, when the two strands of a DNA molecule are separated, each can be used as a pattern or template to produce its complementary strand. The two strands of the DNA is unwound by breaking the weak hydrogen bonds that link the paired bases. They must be held apart to expose the bases so that new nucleotide partners can hydrogen-bond to them. Each template and its new complement together then form a new DNA, identical to the original. The nucleotides are in the center, like the teeth in the zipper, and pair up to hold the two strands together. Importantly, the four different sorts of nucleotides are different shapes, so in order for the strands to close up properly, an A nucleotide must go opposite of a T nucleotide, and a G opposite a C. This exact pairing is called base pairing. When DNA is copied, the two strands of the old DNA are pulled apart by enzymes which move along each of the two single strands pairing up new nucleotide units and then zipping the strands closed. This produces two new pieces of DNA, each containing one strand from the old DNA and one newly made strand.
DNA has a mechanism to proofread the errors done during copying and corrects them through various DNA repair processes. Repair enzymes recognize structural imperfections between improperly paired nucleotides, cutting out the wrong ones and putting the right ones in their place. However, errors are still made when DNA is making a copy of itself because some errors are unavoidable due to the laws of physical chemistry. Nature has developed proofreading techniques that eliminate most errors but cannot correct 100% of the errors. This allows future adaptations to a changing environment. In fact, there is no way to correct 100% of the errors; the correction machinery has some error rate built into it by the laws of physical chemistry such that even if you started with an absolutely perfect copy, the error correction machinery would put an occasional error in the final DNA chain. The language of DNA is extremely complicated, very complex, and yet very precise. However, the estimated average mistake that is not caught turns out to be one error per 10 billion letters. If a mistake occurs in one of the most significant parts of the code, which is in the coding portion of genes, it can cause a disease such as sickle-cell anemia. Yet even the best and most intelligent typist in the world couldn’t come close to making only one mistake per 10 billion letters—far from it. The haploid human genome is estimated to be about 3 billion base pairs (6 billion letters) long. Therefore, the expected ratio of cells making mistakes during copying (DNA replication) is one letter in every two cell divisions. Thus, no matter how precise the DNA replication is, the mutations are inevitable among organisms and become permanent mutations after the next cell division. This is because once such mistakes are established, the cell no longer recognizes them as errors.
Every single living cell has this ability, a chemical machinery to reproduce the DNA from a complementary strand. The controversial part is we have the recombinant DNA technology or DNA cloning in our laboratories to duplicate that machinery out of living cells so we can make complementary strands. The DNA of interest can then be propagated in a foreign host cell. This technology has been around since the 1970s, and propagation has become a common practice in molecular biology labs today. To clone a gene, a DNA fragment containing the gene of interest is isolated from chromosomal DNA using restriction enzymes and then united with a plasmid that has been cut with the same restriction enzymes. Recombinant DNA technology is important for learning about other related technologies, such as gene therapy, genetic engineering of organisms, and sequencing the human genome. Imagine if we had the technology to sequence a pigeon genome to tell us where each gene is located, which is expected very soon. By the way, you might not believe it, but there are human clones among us right now. They weren't made in a lab, though. They are identical twins, created naturally, but we also have the technology to create them artificially.
When cells reach a certain size, they usually either stop growing or divide. Not all cells divide; some, such as nerve, skeletal muscle, and red blood cells, do not normally divide once are mature. Other cells undergo a sequence of activities required for growth and cell division. The stages through which a cell passes from one cell division to the next are referred as the cell cycle. Timing of the cell cycle varies widely, but in actively growing plant and animal cells, it is about 8 to 20 hours. Much of a cell’s life is spent in interphase, the time when no cell division is occurring.
Why study pigeon genetics?
The biggest problem with pigeon genetics is there are not many people who truly understand even the basics. In fact, there are many gross misunderstandings and just plain false beliefs among pigeon breeders, most of whom are badly confused. You might be wondering as a pigeon breeder, why did you have to read about genes, chromosomes, DNA, wild-type, and mutation, etc? Let’s use homing pigeons as an example. A homing pigeon can fly faster than most other breeds and find its home from miles away because it has highly specialized genes that enables such a unique and distinctive ability. The homing pigeon’s intelligence, genetics, and physical ability were specifically designed (selected) to allow it to fly farther and faster than most other breeds, as well as find its way home even from long distances. So, if you understand what genes are responsible for this and how many genes are involved for this breed’s unusual navigational ability, then you will have a better understanding and appreciation for your birds and your hobby. Different pigeon breeds have different mutant abilities or physical characteristics (traits) that make them distinctive, and genes are responsible for this.
As pigeon breeders, we can make genetic changes happen much faster than nature simply because we can select traits more rigorously than nature. We can also select related birds to mate, while in nature related birds very seldom are going to mate. Therefore, both selection and inbreeding speeds the process dramatically. In any species, there will usually be several variants, or alleles (alternatives), of each gene. The alleles of a specific gene are related to each other -- new alleles arise from older ones by mutation or transcription errors (mistakes in copying). Thus, all the pigeons have the same number of genes in them because they all represent the characteristics of one species. However, just like every organism, each pigeon has the different versions of the genes from their possible gene pool.
 What
about the chromosomes? Unlike male pigeons, female pigeons only have
one functional sex-chromosome. This is extremely significant because
there are a lot of genes, including the number of plumage colors that
are located in the sex-chromosomes. If we consider the total number
of chromosomes in a pigeon's DNA, a reasonable estimate would be about
5% of all genes are located on the sex-chromosome, so they are not
particularly important from a genetics standpoint. However, knowing
which genes are located on the sex-chromosomes and which genes are
located in the autosomal chromosomes could help pigeon breeders with
the expected mating results. This knowledge will help breeders to notice
the pedigree mistakes done by others. I have seen some pedigrees where
a red check was supposedly produced from two blue parents; anyone who
understands some basic genetics would know that should not have happened.
These matings are known as sex-linked matings, a mating in which sex
linkage of a particular gene is used to determine the sex of an offspring
and where we can predict the possible colors and the gender of the
offspring. Understanding how many genes that directly affect the visible
traits are located on the sex chromosome can help breeders understand
the reasons for expected plumage color.
What
about the chromosomes? Unlike male pigeons, female pigeons only have
one functional sex-chromosome. This is extremely significant because
there are a lot of genes, including the number of plumage colors that
are located in the sex-chromosomes. If we consider the total number
of chromosomes in a pigeon's DNA, a reasonable estimate would be about
5% of all genes are located on the sex-chromosome, so they are not
particularly important from a genetics standpoint. However, knowing
which genes are located on the sex-chromosomes and which genes are
located in the autosomal chromosomes could help pigeon breeders with
the expected mating results. This knowledge will help breeders to notice
the pedigree mistakes done by others. I have seen some pedigrees where
a red check was supposedly produced from two blue parents; anyone who
understands some basic genetics would know that should not have happened.
These matings are known as sex-linked matings, a mating in which sex
linkage of a particular gene is used to determine the sex of an offspring
and where we can predict the possible colors and the gender of the
offspring. Understanding how many genes that directly affect the visible
traits are located on the sex chromosome can help breeders understand
the reasons for expected plumage color.
Knowing what DNA is, is also very important, especially when the cell divides and duplicates itself. We know that cells have this extraordinary ability to duplicate themselves, and they have proofreaders to check for the errors after the copying process. Yet, errors still happen, and these mutations are the basis of evolution. As pigeon breeders, we need to understand this phenomenon because all of our domesticated breeds are considered to be mutants from the wild-type, the rock dove, scientific name “Columba livia” (our standard). The reasons I gave above are only a few examples of why we should be familiar with basic biological and genetic terms and concepts, but understand that I only barely covered what they are and what are they able to do as scientists are still working diligently to find out more about them.
When you read the remaining parts of these pages, you will understand even more why studying genetics can be very important in your successful breeding program. For instance, you could have a desired bird in your stock, and you could try reproducing that factor without realizing that the desired characteristic is the result produced by more than one gene. Most traits are produced by multiple genes. Moreover, any serious breeder should be familiar with the wild-type concept as a “standard” to gain knowledge of each gene's affect on wild-type, and how to unite the desired traits in one individual. Without this knowledge, you will waste your time with trial-and-error breeding methods, which offers you nothing more than random hope. If you continue reading and understanding the distinction between genotype and phenotype, you will be able to manage your stock better and would not cull the birds which, in fact, could be your foundation birds. Just because a bird does not display any desirable characteristics doesn’t mean it doesn’t carry the necessary genes in its genetic makeup. So don’t feel as though once you start reading something, you must finish it. If you are a beginner, just the information you have read above is too much to digest all at one time. Take a break when you must, but I promise you if you came this far, your genetic knowledge is already way above and beyond most pigeon breeders out there.
 Dominant and
Recessive Alleles
Dominant and
Recessive Alleles
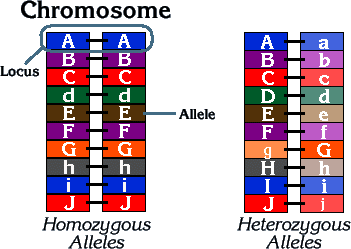
It is important to understand that with some exceptions, chromosomes occur in pairs. Therefore, each gene has more than one form of expression, a matching copy at the same location on its neighboring chromosome. For instance, an eye color gene can have two different forms of expression of orange or pearl eyes in pigeons. These forms of expressions are called alleles or allelomorphs. Each pair of alleles generally has a dominant allele and a recessive allele. In the presence of a dominant allele, the recessive allele does not express itself. The only time that a recessive trait will be seen is when both of the recessive alleles for that trait are present in the genome (a//a or b//b). In genetics, dominance describes a relationship between different forms (alleles) of a gene at a particular physical location (locus) on a chromosome. Alleles may occur in pairs, or there may be multiple alleles affecting the expression of a particular trait. In complex species like humans and pigeons, most traits are determined by more than two alleles. Multiple forms of the allele may exist (like the pattern series in pigeons or the eye colors in humans) though only two will attach to the designated gene site. When expressing dominant and recessive alleles, the dominant allele is always written as a capitalized letter, and the recessive allele is written using the same letter, but in lower case.
When the two alleles at the same gene locus are identical, the individual is called a homozygote for that trait and is said to be homozygous (AA or dd); if instead the two alleles are different, the individual is a heterozygote for that trait and said to be heterozygous (Aa or Bb). The corollary is that homozygous can produce only one kind of gamete and, therefore, breed true, while heterozygous segregate alleles and, for one gene pair, produce two kinds of gametes. When the trait is homozygous, the phenotype is what both alleles represent. If it is heterozygous, then the two alleles interact, and the dominant one is the one that is expressed. Hemizygous gene has no allelic counterpart or is present as only a single copy instead of the usual two copies in a diploid cell or organism. In other words, a gene wherein one of its pair is deleted is a hemizygous gene. Most of the genes of the X chromosome and Y chromosome in human males are hemizygous since males have only one X chromosome (and one Y chromosome, unlike females that have two X chromosomes).
Let me try to explain the concept of alleles and dominance, which is very important to understand when studying genetics, by explaining the human hair and eye colors. Scientists believe that people’s hair color contains four alleles: black is dominant to brown; brown is dominant to blond, and blond is dominant to red. Therefore, blond hair can be a dominant and a recessive depending on which allele it’s paired with. It is very important to understand that out of four possible alternatives (alleles), every human carries only two of the possible hair pigment alleles, which are donated by each parent. You might ask why there are so many different variations (colors) of each possible pigment. Why doesn’t all brown hair look the same, for instance? Because there are other color-modifying genes in every human that makes every brown pigment appear different.
People with red hair must have two red hair alleles (homozygous), and that is the only way they can have (show) red hair. A person with brown hair definitely has one brown allele, but the other allele can be brown, blond, or red. Since brown is dominant to blond and red, these alleles can be carried but cannot present themselves in the presence of a more dominant allele (brown in this case), and therefore the hair color of this person will be brown. However, a person showing brown hair cannot have a black allele because black is dominant to brown. People with black hair can be carrying another black (homozygous for black), brown, blond, or red allele as an alternate to the black they currently possess and show. This is why red hair can show up in anyone’s family in any generation. Both parents pass the red allele that they are secretly carrying on to their child. This could create a lot of confusion for people when their child has red hair, yet the parents both have jet-black hair, but it is genetically possible.
 Our
current understanding suggests that there is one major gene that determines
the hair color in humans, and it works with some modifier genes. Eyes,
on the other hand, are still not understood well because we don’t
exactly know in which chromosome they are located yet. In addition,
eye color in humans is more complicated because there's more than one
locus that contributes to the pigment production in eyes. What we do
know is that black is dominant to brown, which is dominant to blue,
which is dominant to green, which is dominant to gray. Gray eyes are
like red hair in humans, where both alleles have to be gray in order
for a person to possess gray eyes. Eye color is determined by the amount
of a pigment called melanin that is present
in the iris of the eye. Now, the quantity this pigment produces is
determined by the gene’s
controlling pigment production. We know that brown eyes have a good
amount of this pigment, whereas blue eyes have less of it. Geneticists
don’t know how many genes are responsible for eye color yet.
All we know so far is that some genes come in only one or two allelic
versions. Others may come in hundreds of different colors.
Our
current understanding suggests that there is one major gene that determines
the hair color in humans, and it works with some modifier genes. Eyes,
on the other hand, are still not understood well because we don’t
exactly know in which chromosome they are located yet. In addition,
eye color in humans is more complicated because there's more than one
locus that contributes to the pigment production in eyes. What we do
know is that black is dominant to brown, which is dominant to blue,
which is dominant to green, which is dominant to gray. Gray eyes are
like red hair in humans, where both alleles have to be gray in order
for a person to possess gray eyes. Eye color is determined by the amount
of a pigment called melanin that is present
in the iris of the eye. Now, the quantity this pigment produces is
determined by the gene’s
controlling pigment production. We know that brown eyes have a good
amount of this pigment, whereas blue eyes have less of it. Geneticists
don’t know how many genes are responsible for eye color yet.
All we know so far is that some genes come in only one or two allelic
versions. Others may come in hundreds of different colors.
In all populations, children are most commonly born without the pigmented eyes. Some babies have light colored eyes before the age of one. As the child develops, melanocytes (cells found within the iris of human eyes, as well as skin and hair follicles) slowly begin to produce melanin. Because melanocyte cells continually produce pigment, eye color, in theory, can be changed. Most eye changes happen when the infant is around one year old. Observing the iris of an infant from the side, using only transmitted light with no reflection from the back of the iris, it is possible to detect the presence or absence of low levels of melanin. An iris that appears blue under this method of observation is more likely to remain blue as the infant ages. An iris that appears golden contains some melanin even at this early age and is likely to turn green or brown as the infant ages.
One of the most puzzling aspects of eye color genetics is accounting for the birth of brown-eyed children to blue-eyed parents. This is a real phenomenon and not just a case of mistaken fatherhood. Scientists are still trying to solve this puzzle. It seems there are some missing links yet to be discovered in order to understand the exact pattern of the eye color inheritance to be determined genetically. The color of the eye is determined by the amount of a dark brown pigment—melanin—present in the iris of the eye. If the amount is too much, the eye goes brown or even black. If the amount is scarce, the eye turns blue. With in-between amounts, the eye color may go gray, green, hazel, or some lighter shades of brown. If the amount is nil, the eye takes its hue from the blood vessels in the back of the eye and appears to be pink. Albinos are an example of this.
Autosomal & Allosomal
(Sex) Chromosomes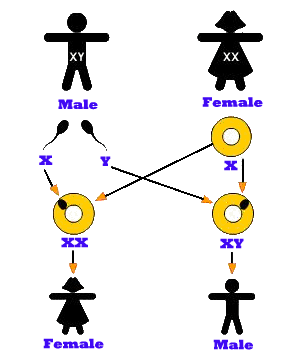
There are two different types of chromosomes. One of them is called an autosome, and the other is called an allosome (sex) chromosome. For example, in humans there are 22 pairs of autosomal chromosomes and 1 pair of sex chromosomes. So in a sense, the autosomal chromosomes has nothing to do with the sex chromosomes. Sex chromosomes determine the offspring's sex, and autosomal is simply all of the other chromosomes. An autosome is a chromosome that is not a sex chromosome – that is to say, there are an equal number of copies of the chromosome in males and females. In humans, there are 22 pairs of autosomes, and additionally, there are X and Y chromosomes which are sex chromosomes. The X and Y chromosomes may be different lengths or have different structure or both. In the picture shown on the right, two chromosomes, X and Y, are used to represent and determine the sex of a human offspring. Male is represented by two distinct sex chromosomes X and Y, and they are called the heterogametic sex. Females have two of the same kind of sex chromosome X and X, and they are called the homogametic sex. In reproduction, each parent submits one chromosome for the child. If the male sperm that joins the egg happens to contain an X chromosome, then it will join with the X of the mother, making the child female (XX). But if the male sperm that joins the egg happens to contain a Y chromosome, the child will be male (XY). In this manner, parental genes decide whether the offspring will be male or female. This also means the gender ratio in humans is 50% male and 50% female. In humans, the male's chromosomes determine the sex of the offspring.
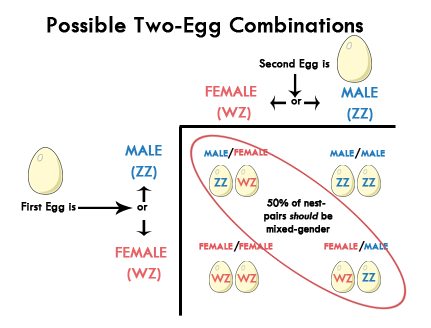
So if we decide to have two children, then there are four possible combinations - boy-boy, girl-girl, boy-girl, and girl-boy. Consequently, two-child pairs will include one boy and one girl, and the other two will be either both boys or both girls. In pigeons, we see the exact opposite of human reproduction. More than 50% of pigeon squab nest-mates are male-female, and there isn’t any scientific evidence to explain this anomaly.
Pigeons have 80 chromosomes arranged in 40 pairs, one of which is the sex chromosomes. But it’s males who have the same chromosome called “Z” - while females bear one “Z” and one “W”. Only female pigeons carry a “W” chromosome. So the chances of a pigeon pair conceiving a male or female squab is, generally speaking, fifty percent. However, studies show that under normal circumstances 70% of the time, the first laid egg is male, and the second laid egg is female. Since newly hatched pigeons are known to double their size in 48 hours, this gives the first-hatch squab greater chance of survival. This brings up another phenomenon if there is equilibrium on male vs. female pigeon numbers in the wild. This is why studying genetics gets interesting where it requires us to be able to organize information, draw conclusions from that information, and find solutions to problems. Even though chances of getting a boy or a girl is 50% in humans, estimated human sex ratio in the world is assumed to be that for every 105 boys there are 100 girls and the ratio fluctuates between 103 to 107 boys for every 100 girls. Thomas W. Pike wrote an excellent article, “Sex Ratio Manipulation in Response to Maternal Condition in Pigeons,” suggesting that mothers in relatively poor physical condition should overproduce daughters by manipulating maternal body condition around the time of egg laying by continuous egg removal and differing feeding regimes. Click here to read this article.
 When
we refer to the female pigeon’s sex-chromosome, we use the term
hemizygous instead of heterozygous as hens have only one mismatched
functional sex-chromosome and no alternative copy for it. That is why
in pigeons, the female's sex-chromosome determines the sex of the offspring
because males are the homogametic sex (ZZ) while females are the heterogametic
sex (ZW).
When
we refer to the female pigeon’s sex-chromosome, we use the term
hemizygous instead of heterozygous as hens have only one mismatched
functional sex-chromosome and no alternative copy for it. That is why
in pigeons, the female's sex-chromosome determines the sex of the offspring
because males are the homogametic sex (ZZ) while females are the heterogametic
sex (ZW).
In insects and mammals the female have two X’s (XX) and the male has one X and one Y (XY). In birds, moths, butterflies, and some amphibians, the sex chromosomes are labeled as Z and W to remind us the switch in these groups, wherein ZZ = male and ZW = female. Thus, in pigeons a cock (male bird) always inherits a Z chromosome from each of his parents, resulting in a sex chromosome set or pair of two Z’s. ZZ is the symbol to represent a cock bird. A hen (female bird), on the other hand, is represented as ZW as she gets Z (functional sex chromosome) from her father and W (non-functional sex chromosome) from her mother, and W has no known gene located on it.
Consequently, in pigeons the sex of the offspring is determined by the Z or W sex chromosome that the female donates during reproduction. Cock birds have two Z chromosomes, and they can only donate one of the Z’s. If the hen also donates a Z, then the offspring will have two Z’s, each donated by a parent, and will make that bird a cock (ZZ) bird. If the hen donates the W sex chromosome to her offspring, then the offspring will have a Z from father and a W from mother and therefore will be a hen (ZW).
Summary
Genetics, the science of genes, heredity, and the variation of organisms, is fundamental to understanding how organisms, populations, and species evolve, behave, and receive common traits from previous generations. A cell is the smallest and most fundamental unit of life that contains the body’s hereditary material and can make copies of itself. Cell numbers increase by cell division, which is where a mother cell divides into two daughter cells. Cell division is an essential feature of life, and we should look at cell division as almost the definition of life.
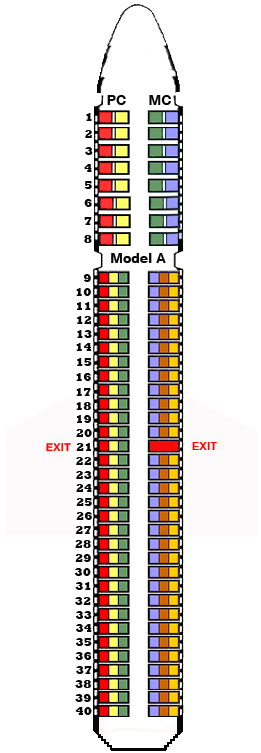 I really
like the analogy given by Ronald Huntley of using a passenger airplane
to explain the basic concepts of genetics for pigeons. Let's
consider the airplane as a pigeon, and let’s assume that all
seats located in rows and in two columns separated by the aisle inside
the airplane make up the DNA of the pigeon. Let’s also assume
that each seat has a long and unique sequence indicating in which row
and in which column they are supposed to be located. Each of these
seat sequences has the same start and end sequences, but the sequence
in between start and end are all unique to each seat, and they all
have different lengths. In this analogy, we will assume these sequences
as the genes. There are 40 rows in this airplane, 40 on the right and
40 on the left. In this analogy, let's consider all rows as the diploid
chromosomes, containing two complete sets of chromosomes, one from
each parent.
I really
like the analogy given by Ronald Huntley of using a passenger airplane
to explain the basic concepts of genetics for pigeons. Let's
consider the airplane as a pigeon, and let’s assume that all
seats located in rows and in two columns separated by the aisle inside
the airplane make up the DNA of the pigeon. Let’s also assume
that each seat has a long and unique sequence indicating in which row
and in which column they are supposed to be located. Each of these
seat sequences has the same start and end sequences, but the sequence
in between start and end are all unique to each seat, and they all
have different lengths. In this analogy, we will assume these sequences
as the genes. There are 40 rows in this airplane, 40 on the right and
40 on the left. In this analogy, let's consider all rows as the diploid
chromosomes, containing two complete sets of chromosomes, one from
each parent.
Just like most passenger airplanes, we have a first-class section. This section has wider but fewer seats in each row. Although most rows have an equal number of seats on each side, one row in the middle, where the exit seats are located, has an unequal number of seats on one side. This particular model is called Model A (female). The other model comes with an equal number of seats in every single row, and it is called Model B (male). In both models, these 40 rows are separated by an aisle. One side is inherited from a bird’s mother (maternal chromosomes - MC), and the other 40 rows, which are perfectly aligned with their counterparts, are inherited from bird’s father (paternal chromosomes - PC). The reason there are first-class seats is that not all chromosomes (rows) and not all genes (seats) are the same length; therefore, some chromosomes are longer and have more genes (seats) in them. Thus, in reality the length of the genes vary greatly in different species, and these unequal bases of genes spread out to all chromosomes randomly.
It is important to note that each seat on the opposite side of each row corresponds with another seat on the opposite side of the same row. For example, in first-class rows 1 through 8, there are red and yellow seats on the PC column (paternal chromosomes) and green and purple seats on the MC column (maternal chromosomes). Therefore, the red seat of the PC column corresponds with the purple seat of the MC column, and the yellow seat of the PC column corresponds with the green seat of the MC column. If these seats, which are located at exactly on the opposite side of each other, have the same exact sequence, we call them a homozygous pair. If one of the sequence is even slightly different, we call them a heterozygous pair and consider them as alleles (alternatives) since their sequence differs from each other. In rows 9 through 40, we have the same concept where each red of the PC column corresponds with each orange of the MC column, each yellow of the PC column with each brown of the MC column, and each green of the PC column with purple of the MC column. In row 21, since the maternal chromosome (W) lacks any genes, that leaves the paternal chromosome (Z) in hemizygous state – a gene that has no allelic counterpart.
Now consider every seat in this scenario as a gene and their assigned location as the locus. Each seat can only have one unique sequence allele of each gene. Recall that on paired chromosomes, every gene has a corresponding opposite gene on the other chromosome. Thus, at the location of a given gene, there can only be two possible alternative genes that can occupy that locus (one loci on each chromosome). There may be several alternative mutant genes that can occupy that location, but only two can occupy at any given instance (except the genes located on the sex chromosomes – row 21 in this analogy). Also note that in reality, chromosomes have thousands of genes, but from our airplane analogy, we only have 2 or 3 seats (locus points) on each row.
In this analogy, consider each row on each side of the aisles as a haploid chromosome, a single set of unpaired chromosomes. Depending on the exit row, this bird is either a cock bird, if it is model B and has an equal number of seats in every row, or it is a hen if it is model A and has one row that has unequally numbered seats. Nonetheless, both model A and model B have 40 total rows separated by an aisle. Recall that pigeons have 40 pairs of chromosomes, half from their mother and half from their father, so pigeons have 80 individual chromosomes (rows in our analogy). That unique row in these particular models represents the sex chromosomes (row 21 in our analogy). Male pigeons have two identical rows with an equal number of seats and passengers (loci and genes), which are represented as (ZZ). Female pigeons, on the other hand, have two distinct exit rows, which are represented as (ZW). The row known as the W has no known seats, hence genes in them.
Recall that each seat has a long sequences of numbers (gene coding, three-letter words made out of A-T-G-C letters), and an attendant (enzyme) at the assembly line checks to make sure that these sequences are matching with the database (wild-type DNA coding). Recall that during cell division, DNA is replicated, and a machinery is designed to catch and correct the mistakes during copying. Unfortunately, this mechanism can’t always catch all the mistakes because they (genes) are very, very long and sometimes copiers make mistakes copying these sequences. However, without these mistakes, we would not have mutations, and the evolution would stop. We would be breeding and flying only blue bar pigeons that had the exact same gene sequencing. The assembly line attendants check to see if every sequence is copied correctly.
At the end, all the sequences are reported to the control center (mechanism to read the gene coding instructions and start ordering necessary items to build the organism). For instance, let’s assume that one of the sequences (allele genes) on the pattern seats (known as the C loci in pigeon genetics) have the correct gene coding producing bar pattern (wild-type - sybolized as + sign), but the other one has spelling mistakes (mutation that can code for either barless, checker, or T-checker—alleles of bar pattern). In this scenario, let’s assume the second seat is coding for the checker pattern (+//C). Let’s also assume that at the indigo loci, we have unmatched seat sequences, and one of them is not wild-type (+//In). At the recessive red loci, one of the locus points has a wild-type coding, so no problems are there. But on the opposite side of the aisle on the same chromosome, the other gene at the recessive red loci is coding for recessive red mutation (+//e). At the eye color loci, one of the alleles is coding for orange (wild-type), and the other allele is coding for the mutated pearl eye color (+//tr). Finally, let’s assume that only one side of the row 21 (sex chromosomes) has seats, and the other side has no seats and therefore non-functional (ZW). Let’s assume that in our analogy, the rest of the genes located in every loci in all other chromosomes are coding for wild-type. Let’s also notice, but ignore it for the sake of argument, that the picture example I am using here is a homing pigeon, and it must have all the necessary genes to affect its body type and enable this pigeon with the necessary traits that a usual homing pigeon would normally have.
 When
these errors (mutations) are reported to the assembly line, instead
of building a blue bar (wild-type) bird based on this blue print, the
order is to build a female hetero indigo, hetero checker homing pigeon
with orange eyes that carries recessive red. Why? Because based on
breeding data, we know indigo is a dominant mutation to wild-type and
does not have any other known alleles. Checker is also a dominant allele
to wild-type (bar) and therefore will present itself when the other
allele present is either wild-type or barless—the only recessive
allele of wild-type bar pattern. The pearl eye color is recessive to
orange eyes (wild-type) and therefore can only express itself in the
homozygous state. So, the pigeon in our analogy will have orange eyes.
When
these errors (mutations) are reported to the assembly line, instead
of building a blue bar (wild-type) bird based on this blue print, the
order is to build a female hetero indigo, hetero checker homing pigeon
with orange eyes that carries recessive red. Why? Because based on
breeding data, we know indigo is a dominant mutation to wild-type and
does not have any other known alleles. Checker is also a dominant allele
to wild-type (bar) and therefore will present itself when the other
allele present is either wild-type or barless—the only recessive
allele of wild-type bar pattern. The pearl eye color is recessive to
orange eyes (wild-type) and therefore can only express itself in the
homozygous state. So, the pigeon in our analogy will have orange eyes.
Recessive red mutation is an autosomal recessive trait and needs two copies of the mutated genes (homozygous state) to present itself. In our example, since the other allele located in recessive red loci was the wild-type, the wild-type gene overwrote the instructions and made this particular bird, not a recessive red. Had the recessive red gene been present in both of the recessive red loci, this bird would have presented itself as recessive red, and we would not see any indication whatsoever of the checker pattern or the indigo coloration. Indigo and checker pattern are both dominant alleles of wild-type, and they only need one copy to present themselves. However, recessive red is epistatic, which would have concealed the check pattern, and any other pigment production responsible producing the indigo colors.
Next time you look at a pigeon, imagine that they are passenger airplanes with 40 rows. If they don't look like our standard blue bar, then they must have some seats (genes) with the mutated sequences in different rows (chromosomes) to make that bird appear different than our standard. We will talk about how each seat got their assigned seat sequences and how each row separated by the aisle has become the combination of both maternal and paternal chromosomes in the next page.